Molecular mechanisms of asymmetric cell division
How can cell division generate daughter cells that differ from one another? This can be achieved by two fundamentally distinct ways of asymmetric cell division. The cell division process can create intrinsically different daughter cells, or alternatively, initially identical daughter cells can be instructed to take different fates dependent on the external signals they receive (Horvitz and Herskowitz, 1992). The plane of cell cleavage is crucial for both forms of asymmetric division. Specifically, the cleavage plane determines the size of daughter cells as well as the distribution of cell fate determinants in intrinsically asymmetric division, and the position of daughter cells within a tissue or with respect to a stem cell niche in extrinsically asymmetric division.
The plane of cell cleavage is crucial for both forms of asymmetric division. Specifically, the cleavage plane determines the size of daughter cells as well as the distribution of cell fate determinants in intrinsically asymmetric division, and the position of daughter cells within a tissue or with respect to a stem cell niche in extrinsically asymmetric division.
The cell cleavage plane is determined by the position of the mitotic spindle, as cleavage takes place perpendicular to and usually midway through the spindle. Hence, the position of the spindle determines if polarized cells divide symmetrically or asymmetrically. When the spindle aligns along the axis of polarity, cell cleavage generates different daughter cells, while spindle alignment perpendicular to the polarity axis allows the formation of equal daughter cells.
What, then, determines the position of the spindle? Our previous studies in C. elegans identified an evolutionarily conserved pathway for spindle positioning. This pathway involves the coiled-coil protein LIN-5, as well as the G protein regulators GPR-1/2 that mediate association of LIN-5 with the alpha subunit of a heterotrimeric G protein. Similar protein interactions have been identified in Drosophila (Mud—Pins—Gα) and mammalian cells (NuMA—LGN—Gα). Membrane associated LIN-5—GPR—Gα recruits a dynein motor complex to the cell cortex. The minus-end-directed motor activity of dynein, combined with microtubule disassembly, is thought to generate pulling forces that move the spindle poles and chromosomes to the cortex. Asymmetries in these cortical pulling forces cause displacement or rotation of the mitotic spindle, and thereby control the plane of cell cleavage.
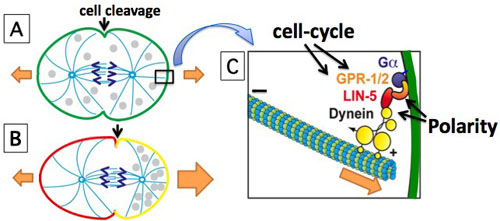
The mitotic spindle (light blue) segregates the chromosomes (purple) and instructs cell cleavage to take place between the spindle poles. (A) Division of unpolarized cells generates equal daughter cells. (B) Polarized cells that divide perpendicular to the polarity axis produce unequal daughter cells. (C) Ga/GPR/LIN-5/dynein generate the forces that pull astral microtubules to the cortex in chromosome segregation and spindle positioning (orange arrows). Cell-cycle and polarity proteins control the timing and location of these pulling forces. Our results indicate that local inhibition of LIN-5 function by anterior PAR/PKC-3 leads to posterior displacement of the spindle and asymmetric cell cleavage.
Recently, we found that the polarity kinase PKC-3 (aPKC, atypical protein kinase C) directly regulates the LIN-5 (NuMA)—GPR (Pins/LGN)—Gα pathway, in order to position the mitotic spindle. We developed a quantitative mass spectrometry approach for in vivo identification of protein kinase substrates. Starting from this method, we demonstrated that anterior located PKC-3 inhibits cortical microtubule pulling forces through direct phosphorylation of LIN-5. This provides an important molecular mechanism by which polarity cues control the spindle-positioning machinery to instruct the cleavage plane during asymmetric cell division.
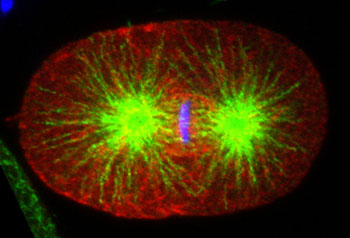
Anterior cortical enrichment of phosphorylated LIN-5. Fluoresence micrograph of wild-type C. elegans embryo at metaphase of the first division, stained with anti-pS737 LIN-5 antibody (red) and anti-tubulin (green). DNA is in blue. Scale bar, 10µm.
Our current efforts focus on resolving how cell-cycle and cell-polarity kinases together regulate the function and position of the spindle apparatus in space and time, in order to coordinate chromosome segregation with cell fate determination. Using the lessons from the one-cell embryo as a starting point, we have initiated studies of asymmetric cell division in more complex systems, in particular stem cell divisions in the C. elegans epidermis.
Back to Overview